1. Introduction
In our books species are described in a standardized manner, thus making it easy to compare different species and also to find pertinent information about them.
Genera and species are placed alphabetically without regard to relationships as this should make the books easier to use. The taxonomy of the group is treated in a separate chapter.
It is inevitable that mistakes have crept in and that there are omissions. We will of course be grateful to be informed about such cases and to have suggestions on improvements. The author’s addresses are given above.
The distribution of species is, in most cases, given in a general way since our knowledge about the distribution for most species, is very scanty and accidental. Still many countries and areas are more or less mycologically unexplored due to lack of trained mycologists.
What is a polypore? ‑ definition of the group.
Linnaeus, the father of plant systematics, used Boletus as the genus for all fungi with tubes or pores whether the basidiocarps were fleshy and stalked or woody and sessile. Soon however, the concept changed and Fries (1821), in his basic work Systema Mycologicum, clearly separated the polypores as a group of its own in a "Tribus Polypori" although he included some genera which today are included in other taxa.
The family name Polyporaceae was first used by Fries in 1838 and denoted most of the polypores although excluding some genera with a lamellate hymenophore such as Lenzites, which he placed among the agarics. Later, Polyporaceae was used somewhat vaguely, especially in floras and floristic lists, to include all types of fungi with pores but also other types of hymenophore but clearly related to the "true" polypores.
In this mycota we have tried to include all fungi that would naturally be sought in a manual of polypores although we clearly acknowledge that this is a mixed group of families, with different phylogenetic backgrounds. The reader is again referred to the chapter on Taxonomy for more information on the classification of polypores included in this flora.
In Corticiaceae (s. lato) there are a restricted number of species with a distinctly poroid hymenophore – these are included here.
An introduction to poroide fungi
All introduction chapters:
1: Introduction
9. Practical advice for collection and determination of polypores
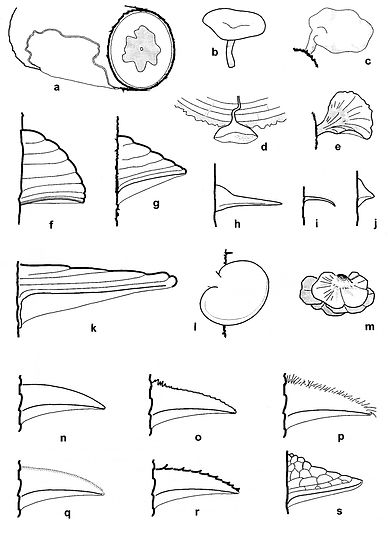
Fig. 1. Types of basidiocarps, a) resupinate, b) centrally stipitate, c) laterally stipitate, d) pendant, e) spathulate/fan shaped f) ungulate g) triquetros, h) umbonate, i) deflexed, j) nodulose, k) applanate, l dimidiate, m) imbricate, n) glabrous, o) scrupose, p) hirsute, q) velutinate, r) hispid, s) rimose. Del. T. Niemelä.